
চোখের বিবর্তন (The evolution of Eye) শেষ অংশ
রিসেপ্টরের আবির্ভাব:
ট্রেভর ল্যাম্বের টীম যখন গবেষনা করছিলেন তিন স্তর বিশিষ্ট রেটিনার বিবর্তন নিয়ে, চোখের বিবর্তন সংক্রান্ত আরেকটি প্রশ্নর মুখোমুখি হতে হয়েছে তাদের। তা হচ্ছে ফটোরিসেপ্টর বা আলোক সংবেদী কোষের বিবর্তন। প্রানীজগতে ফটোরিসেপ্টর কোষগুলো সাধারণত সুনির্দিষ্ট দুইটি শ্রেনীতে ভাগ করা সম্ভব: রাবডোমেরিক (Rhabdomeric) এবং সিলিয়ারী (Ciliary)। অতি সাম্প্রতিককাল পর্যন্ত, বহু বিজ্ঞানীরা মনে করতেন অমেরুদন্ডী প্রানীরা রাবডোমেরিক শ্রেনীর রিসেপ্টর ব্যাবহার করে এবং মেরুদন্ডী প্রানীরা ব্যবহার করে সিলিয়ারী ধরনের ফটোরিসেপ্টর। কিন্তু বাস্তবে ব্যাপারটা খানিকটা জটিল। বহু প্রানীদের মধ্যে দেখা যায় তারা সিলিয়ারী রিসেপ্টর ব্যাবহার করছে দৃষ্টি ছাড়া অন্যান্য কাজের জন্য আলো শনাক্ত করতে; যেমন, সারকাডিয়ার রিদমকে নিয়ন্ত্রন করতে, কিন্তু রাবডোমেরিক রিসেপ্টর আলো শনাক্ত করে শুধুমাত্র দৃষ্টি ক্ষমতার সহায়তার বা কোন কিছু দেখার জন্য। আর্থ্রোপডদের যৌগিক চোখ আর মোলাস্ক, যেমন অক্টোপাসদের ক্যামেরা স্টাইল চোখ, যারা মেরুদন্ডী প্রানীদের ক্যামেরা স্টাইল চোখ থেকে সম্পুর্ণ আলাদা ভাবে বিবর্তিত হয়েছে, তারাও রাবডোমেরিক রিসেপ্টর ব্যবহার করে। কিন্তু মেরুদন্ডী প্রাণীদের চোখ সিলিয়ারী ক্লাসের রিসেপ্টর ব্যাবহার করে তাদের দৃষ্টি অনুভুতি ও দৃষ্টি ক্ষমতার জন্য।
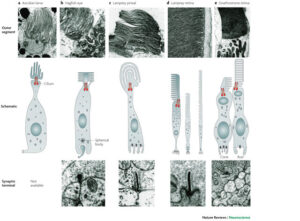
ছবি: উপরের ছবিটিতে দেখানো হয়েছে কর্ডেট এবং মেরুদন্ডী প্রানীদের বিবর্তনের বিভিন্ন পর্যায়ে সিলিয়ারী ফটোরিসেপ্টরদের গঠন: ছবির মাঝের স্তরে দেখানো হয়েছে পুরো সিলিয়ারী রিসেপ্টরগুলোর স্কিমাটিক ডায়াগ্রাম; উপরে এবং নীচের স্তরে দেখানো হয়েছে যথাক্রমে রিসেপ্টরের বাইরের সেগমেন্টের ও সাইন্যাপটিক বা ভিতরের স্নায়ু সংযোগ স্থানের ইলেক্ট্রন মাইক্রোগ্রাফ। ভালো করে লক্ষ্য করলে বোঝা যাবে ধীরে ধীরে বাইরের সেগমেন্টে কেমন করে ল্যামিনার বা পর্দার মতো আরো সুসংগঠিত হয়েছে, আর সাইনাপটিক টার্মিনালে সাইন্যাপটিক রিবনের আবির্ভাব হয়েছে। সুত্র: ট্রেভর ল্যাম্ব ও সহযোগীরা : Nature Reviews: neuroscience; volume 8, December 2007)
২০০৩ এ হাইডেলবার্গ এ ইউরোপিয়ান মলিক্যুলার বায়োলজী ল্যাবে ডেলটেভ আরেন্দ প্রথম রিপোর্ট করেন যে মেরুদন্ডী প্রানীরা তাদের রেটিনায় রাবডোমেরিক ফটোরিসেপ্টরের বংশধর রিসেপ্টর এখনও ধারন করে। অবশ্য এই বংশধর রিসেপ্টরগুলো বেশ বড় আকারের কিছু পরিবর্তনের মধ্য দিয়ে পরিবর্তিত হয়ে রুপান্তরিত হয় আউটপুট নিউরোনে, যাদের কাজ রেটিনা থেকে ব্রেনে স্নায়ু বা তথ্য সংকেত পাঠানো। এই আবিষ্কারের অর্থ হচ্ছে আমাদের চোখের রেটিনায় দুই ধরনেরই ফটোরিসেপ্টরে বংশধর রিসেপ্টর বিদ্যমান: সিলিয়ারী ক্লাস, যা সবসময়ই ফটোরিসেপ্টর এবং রাবডোমেরিক ক্লাস, যা পরিবর্তিত হয় আউটপুট নিউরোনে। বিদ্যমান কোন কিছুকে ( যেমন কোন প্রোটিন বা কোন কোষ ) চাপে ফেলে অন্য নতুন কোন উদ্দেশ্যে ব্যবহার করানোর এই প্রক্রিয়া হচ্ছে বিবর্তনের কাজ করার অন্যতম প্রধান একটা পদ্ধতি। সে কারনেই, অমেরুদন্ডী প্রানীদের থেকে সিলিয়ারী আর রাবডোমেরিক ফটোরিসেপ্টরগুলো আমাদের চোখে যে ভিন্ন দায়িত্ব পালন করে এই আবিষ্কারটি আরো জোরালো প্রমান জোগায় যে, মেরুদন্ডী প্রানীদের চোখের আসলে সৃষ্টি হয়েছে সম্পুর্ন প্রাকৃতিক একটি প্রক্রিয়ায়। গবেষনার স্বার্থেই যদিও আমরা অবশ্য ভাবতে পারি, ঠিক কোন ধরনের পরিবেশের নির্বাচনী চাপ বাধ্য করেছে এই কোষগুলোতে ভিন্ন নতুন দায়িত্ব পালনের পথে বিবর্তিত হতে।
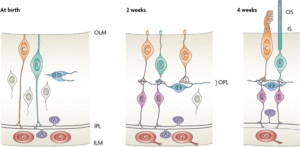
ছবি: উপরের ছবিতে স্তন্যপায়ী প্রানীদের রেটিনায় নিউরোনাল বা স্নায়বিক সংযোগ বা সার্কিট্রির ডেভলপমেন্ট: উপরের ছবিতে দেখানো হয়েছে কিভাবে স্তন্যপায়ী প্রানীদের রেটিনার নিউরাল সার্কিট্রিটা গড়ে ওঠে। এই গবেষনাটি হয়েছে ফেরেটদের চোখে। ফেরেটরা এই গবেষনার জন্য সুবিধাজনক কারন ফেরেটদের জন্ম হয় খুবই অপরিনত অবস্থায়; জন্মের দুই সপ্তাহ পরে কেবল মাত্র এরা চোখ খুলতে পারে। ছবিতে বায়ের প্যানেলে জন্মের সময়, কোন ও রড ফটোরিসেপ্টর (C এবং R) , যাদের আলাদাভাবে চেনা যায়না শুধুমাত্র আকার বা অবস্থান দেখে, সাধারনতঃ এ পর্যায়ে এরা উভয়ই বাইপোলার আকৃতির হয়ে থাকে । কিন্তু রোডোপসিন বা অন্য কোন আনবিক মার্কার দিয়ে এদের পৃথক করা সম্ভব। উল্লেখযোগ্য বিষয়টি হচ্ছে ফটোরিসেপ্টর কোষগুলো তাদের অ্যাক্সোন বা প্রসেস সরাসরি রেটিনার আউটপুট লেয়ারের স্নায়ু কোষের সাথে সংযোগ করে ( এই ডায়াগ্রামের নীচের দিকে লক্ষ্য করতে হবে) বা যাকে ইনার প্লেক্সিফর্ম লেয়ার (IPL) বলা হয়। অবশ্য এখনও অমীমাংসিত আছে, তারা যে প্রথম নিউরনগুলোর সাথে সংযোগ সৃষ্টি করে তারা কি অ্যামাক্রিন কোষ (A) নাকি গ্যাঙ্গলিয়ণ কোষ (G); এ পর্যায়ে হরিজন্টাল সেল (H) এবং বাইপোলার সেল খুব সামান্যই ডিফারেন্টসিয়েটেড বা পরিবর্তিত হয়। মাঝের প্যানেলটি আরো ২ সপ্তাহ পরের রেটিনার অবস্থা , যে সময় ফেরেটরা প্রথম চোখ খোলে, হরিজন্টাল সেল (H) ও বাইপোলার সেল (B) পুর্ণতা পায় এবং ফটোরিসেপ্টরগুলো তাদের প্রসেসগুলো পেছনে সরিয়ে আনে , আগে যারা আউটপুট নিউরোনের সাথে সংযোগ করেছিল, তারাই হরিজন্টাল সেল (H) ও বাইপোলার সেল (B) দের সাথে নতুন করে সংযোগ স্থাপন করে। এই সংযোগ স্তরটার নাম OPL বা আউটার প্লেক্সিফর্ম লেয়ার। একেবারে ডান প্যানেলে আরো পরবর্তীতে (৪ সপ্তাহ) ফটোরিসেপ্টরগুলো আরো পরিবর্তিত হয়ে আলোক সংবেদী আউটার বা বাইরের সেগমেন্ট (OS) ও ইনার বা ভিতরের সেগমেন্ট তৈরী করে (IS); একমাত্র এই পর্যায়ে দেখার মুল সংকেত প্রবাহ মেকানিজমটি কাজ করা শুরু করে। করে। সুত্র: ট্রেভর ল্যাম্ব ও সহযোগীরা : Evolution: Education and Outreach (1: 2008)।
মেরুদন্ডী প্রানীদের রেটিনায় আলোক সংবেদী কোষ হিসাবে কেন সিলিয়ারী ফটোরিসেপ্টরগুলো সফল হয়েছিল, আর রাবডোমেরিক শ্রেনী ফটোরিসেপ্টরগুলো রুপান্তরিত হয়েছে প্রোজেক্শান বা আইটপুট নিউরণে, সেই প্রশ্নটার উত্তর খোজার জন্য ট্রেভর ল্যাম্ব এই সব কোষের মধ্যে বিদ্যমান আলোক সংবেদী পিগমেন্ট বা রোডোপসিন অর্থাৎ যে অপসিন প্রোটিন অনু তারা বহন করে, তা বিশ্লেষন করেছিলেন। ২০০৪ সালে জাপানের কিয়োটো বিশ্ববিদ্যালয়ের ইয়োশিনোরী শিচিদা ও তার সহযোগীরা তাদের একটি গবেষনায় দেখেছিলেন যে, মেরুদন্ডী প্রানীদের ভিসুয়াল (আলোক সংবেদী) পিগমেন্ট এর বিবর্তনের শুরুর দিকে, একটি গুরুত্বপুর্ণ পরিবর্তন হয়, যার ফলে আলোক কর্তৃক সক্রিয় হওয়া বা লাইট অ্যাকটিভেটেড পিগমেন্ট অনুটি আরো খানিকটা বেশী স্থায়ীত্ব ও সেকারনে বেশী সক্রিয়তা লাভ করে। এখানে ট্রেভর ডি. ল্যাম্ব প্রস্তাব করেছিলেন সেই একই পরিবর্তনটি আরো একটি কাজ করেছিল, সেটা হলো সক্রিয় হয়ে ওঠা রোডোপসিন অনুকে পুনরায় আগের নিষ্ক্রিয় অবস্থায় পুনঃ পত্যাবর্তন প্রক্রিয়াটিকে আটকে দেয়া; আবার এই নিস্ক্রিয় অবস্থায় ফিরে আসাটা রাবডোমেরিক রোডোপসিনের জন্য বিশেষ গুরুত্বপুর্ন কারন, দ্বিতীয়বার আলোর ফোটন কনা শোষন করার জন্য তার আগের অবস্থায় ফেরত আসতে হয়; এবং যেটি আবার একটি বায়োকেমিক্যাল প্রক্রিয়া শুরু করার জন্য জরুরী, রাবডোমেরিক রোডোপসিন অনুকে নতুন করে আলোর সংকেত ধারন করার জন্য পুনরায় প্রস্তুত করতে যা একান্ত প্রয়োজন। যখনই এই দুটি ধাপ ঠিক জায়গা মত তাদের কাজ শুরু করে, ট্রেভর ডি, ল্যাম্ব তার হাইপোথেসিসে প্রস্তাব করেন যে, এর ফলে সিলিয়ারী ফটোরিসেপ্টরগুলো রাবডোমেরিক ফটোরিসেপ্টরদের থেকে একটি সুস্পষ্ট বাড়তি একটি সুবিধা পায়, বিশেষ করে সেই সময়ের গভীর সাগর তলদেশের পরিবেশে, যেখানে আলোর পরিমান তুলনামুলক ভাবে বেশ কম ছিল। এর ফলশ্রুতিতে কিছু আদি কর্ডেট ( মেরুদন্ডীদের আদি প্রানীরা ) এ জায়গাগুলোতে বসতি স্থাপন করতে পেরেছিল, রাবডোমেরিক ফটোরিসেপ্টর উপর নির্ভর করা প্রানীদের ক্ষেত্রে যে জায়গায় বসতি স্থাপন করার ক্ষমতা ছিল না -এর কারন তাহলে শুধু উন্নত সিলিয়ারী ফটোরিসেপ্টরের অপসিন বেশী ভালো দেখতে সহায়তা করেছিল তা কিন্তু না ( কারন উন্নত দৃষ্টি ক্ষমতার জন্যে ক্যামেরা স্টাইল চোখের অন্য অংশগুলো এসেছে আরো অনেক পরে), বরং এর কারন হিসাবে বলা যেতে পারে, এটি প্রানীদেহের সারকাডিয়ার রিদম এবং ঋতু নির্ভর চক্রগুলোকে সময়ের হিসাব রাখার জন্য প্রয়োজনীয় আলো শনাক্ত করার সিস্টেমটাকে আরো উন্নত করেছিল।
যেহেতু এই সব আদি কর্ডেটদের বসবাস ছিল অপেক্ষাকৃত অন্ধকার এক জগতে, সিলিয়ারী ফটোরিসেপ্টরের সাথে তাদের বাড়তি অপেক্ষাকৃত কম আলোক সংবেদী যে রাবডোমেরিক রিসেপ্টরগুলো ছিল, তারা মুলত: ছিল এ জায়গার জন্য অপ্রয়োজনীয়, সেকারনেই তারা নতুন কোন দায়িত্ব নেবার জন্য ছিল মুক্ত: ফলে এরা রুপান্তরিত হয়েছিল নিউরোনে, রিসেপ্টর আর ব্রেনের মধ্যে সংকেত আদান প্রদান করার দায়িত্ব নিতে ( এ পর্যায়ে তাদের প্রয়োজন ছিল না কোন অপসিনের, তাই প্রাকৃতিক নির্বাচন এই কোষগুলো থেকে অপসিনকেও সরিয়ে দেয়)।
এবং চোখের জন্ম:
ট্রেভর ডি ল্যাম্ব ও সহযোগীরা যখন ধারনা পেলেন কেমন করে মেরুদন্ডী প্রানীদের রেটিনা নানা অংশের উদ্ভব হয়েছে; এবার তারা জানতে চাইলেন কেমন করে একটি দৃষ্টিক্ষমতাহীন আলোক সংবেদী অঙ্গ থেকে চোখ দৃষ্টি ক্ষমতা সম্পন্ন একটি অঙ্গে পরিনত হয়েছিল প্রায় ৫০০ মিলিয়ন বছর আগে। এবারও তারা তাদের সেই প্রশ্নের উত্তরের ক্লু খুজে পান বিকাশমান ভ্রুণে। ভ্রুণ বিকাশের একেবারে শুরুর দিকে স্নায়ুতন্ত্রের যে অংশটি চোখের জন্ম দেয়, তারা প্রথমে ভ্রুনের সামনের দিকে দুটি স্ফীতকায় থলের মত অংশের সৃষ্টি করে ( যা বলে ভেসিকল); এরপর এই ভেসিকল দুটি আলাদা আলাদা ভাবে ভেতরের দিকে ভাজ হয়ে একটি C আকৃতির রেটিনার সৃষ্টি করে যা চোখের একেবারে ভিতরের স্তরটি তৈরী করে। বিবর্তন প্রক্রিয়াটি সম্ভবত অনেকটা এভাবেই সংঘটিত হয়েছিল বলে ধারনা করেন ট্রেভর ডি. ল্যাম্ব ও সহযোগীরা; তাদের প্রস্তাবনা অনুসারে, এ রকম কোন C আকৃতির দ্বি স্তরের রেটিনা -যার বাইরের দিকটি তৈরী সিলিয়ারী ফটোরিসেপ্টর আর ভেতরের দিকে থাকে রাবডোমেরিক ফটোরিসেপ্টর থেকে রুপান্তরিত হওয়া বর্হিমুখি বা আউটপুট নিউরণ- বিশিষ্ট এধরনের কোন একটি প্রোটো আই বা আদি চোখ -বিবর্তিত হয়েছিল আদি কোন মেরুদন্ডী প্রানীদের পুর্বপুরুষ প্রানীদের শরীরে প্রায় ৫৫০ থেকে ৫০০ মিলিয়ন বছর আগে মুলতঃ শরীরের আভ্যন্তরীন ছন্দ বা সারকাডিয়ান রিদমকে নিয়ন্ত্রন করার জন্য; এছাড়া ধারনা করা হয় এই আদি-চোখ ছায়া শনাক্ত এবং শরীরের অবস্থান ঠিক বা দিক ওরিয়েন্ট করতেও সাহায্য করতো।
ভ্রুণ বিকাশে সময় এর পরের ধাপে রেটিনা যখন আবার ভিতরের দিকে ক্রমশ ভাজ খেয়ে ভেতরের দিকে ঢুকতে থাকে, তখন লেন্স তৈরী শুরু হয়; চোখের লেন্স তৈরী হয় ভ্রুণের শরীরের বাইরের স্তর বা এক্টোডার্ম থেকে : C আকৃতির রেটিনার সৃষ্টি করা বাকানো শুন্যস্থানটায় এক্টোডার্ম এর যে অংশটা স্ফীত হয়ে ঢুকে পড়ে, সেখান থেকে। এই ভিতর দিকে বের হয়ে আসা এক্টোডার্মের বাড়তি অংশটি একসময় বাকী এক্টোডার্ম থেকে সম্পুর্ন পৃথক হয়ে, মুক্ত লেন্সের আকার ধারন করে। ট্রেভর ডি. ল্যাম্ব ও তার সহযোগী ও অন্যান্য বিজ্ঞানীদের ধারনা এরকমই প্রায় সদৃশ কোন একটি পর্যায়ক্রমিক প্রক্রিয়া বিবর্তনের সময়েও ঘটে। যদিও তারা এখনও সঠিক জানেন না ঠিক কখন এই পরিবর্তনটা আসলে ঘটেছিল, কিন্ত ১৯৯৪ সালে সুইডেনের লুন্ড বিশ্ববিদ্যালয়ের গবেষকরা দেখিয়েছেন চোখের অপটিক্যাল অংশগুলো খুব সহজে বিবর্তিত হতে পারে ১ মিলিয়ন বছর সময়ের ব্যাপ্তিতে। আর তাই যদি হয়, কোন দৃষ্টিশক্তিবিহীণ প্রোটো আই থেকে এই ইমেজ বা ছবি সৃষ্টি করার বা দৃষ্টি ক্ষমতা সম্পন্ন চোখ বিবর্তিত হয়েছে ভুতাত্ত্বিক সময়ের প্রেক্ষিতে খুবই অল্প সময়ের মধ্যে।
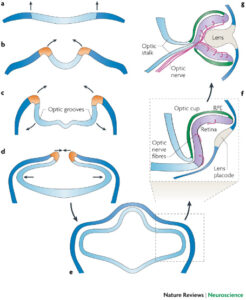
ছবি: উপরের ছবিতে মেরুদন্ডী প্রানীদের আই ক্যাপের ডেভেলপমেন্ট : a) মেরুদন্ডী প্রানীদের নিউরাল প্লেটই হলো নতুন আই ক্যাপ তৈরীর শুরুর জায়গা। b) নিউরাল প্লেটটি উপরে এবং ভিতরের দিকে ভাজ হতে থাকে c) অপটিক গ্রুভের আবির্ভাব হয়, যা ভিতরের দিকে ঢুকতে থাকে d) নিউরাল ফোল্ডের প্রান্ত গুলো কাছাকাছি আসতে শুরু করে আর অপটিক ভেসাইকল বাইরে দিকে আরো বাড়তে থাকে e) প্রান্ত বা ফোল্ডের ঠোটগুলো বন্ধ হয়ে যাবার পর নিউরিাল টিউবের এ অংশটি আলাদা হয়ে যায়। এই পর্যায়ে সামনের ব্রেণের অংশ বা ফোরব্রেন, উপরের দিকে বাড়তে থাকে এবং অপটিক ভেসাইকল বাইরের দিকে স্ফীত হওয়া অব্যাহত থাকে। যখন ভেসাইকল এক্টোডার্ম এর কাছে আসে, তারা তখন লেন্স প্ল্যাকোড তৈরী করতে প্রয়োজনীয় স্টিমুলাস বা প্রনোদনা প্রদান করে f) অপটিক ভেসিকল এর পর নিজের ভিতরের দিকে আবার ভাজ হয়, যার ফলে ভবিষ্যত রেটিনাল পিগমেন্ট এপিথেলিয়াম আর রেটিনা পাশাপাশি চলে আসে, এবং এই দুই স্তরের মাঝে ফাকা জায়গাটা ভেন্ট্রিকলটি একসময় বিলুপ্ত হয়ে যায়। রেটিনাল গ্যাঙ্গলিয়ন কোষগুলো তাদের অ্যাক্সনাল প্রসেসগুলো সারা রেটিনার উপর দিয়ে বিস্তৃত হতে শুরু করে। ভ্রুণের বাইরের পৃষ্ঠে যেখানে এক্টোডার্মের লেন্স প্ল্যাকোডটি আছে, সেখানেই লেন্স পিট তৈরী হয় ((এই ডায়াগ্রামটি ডান চোখের ঠিক মাঝ বরাবর ক্রশ সেকশান করয়েড টিস্যুর মধ্য দিয়ে, সেকারনে শুধু রেটিনার উপরের অংশ আর রেটিনাল পিগমেন্টেড এপিথেলিয়াম (RPE) স্তরটি শুধু দেখা যাচ্ছে)) g) অপটিক ক্যাপটি ধীরে ধীরে পরিধিতে বাড়তে থাকে। এবং একসময় করয়েড ফিসারের মুখটা বন্ধ করে অপটিক নার্ভ এর অ্যাক্সনগুলো বা ফাইবারগুলোকে ঘিরে ধরে (হ্যায়ালয়েড ও রেটিনার রক্ত নালী সহ; যা ছবিতে দেখানো হয়নি। বাইরের স্তর বা এক্টোডার্মাল টিস্যুগুলো আরো বিশেষায়িত হতে থাকে এবং একসময় লেন্স তৈরী করে। সুত্র: ট্রেভর ল্যাম্ব ও সহযোগীরা : Nature Reviews: neuroscience; volume 8, December 2007)
আলো সংগ্রহ এবং ছবি (ইমেজ) ফোকাস করার লেন্স আবির্ভাব হবার সাথে সাথে চোখে দৃষ্টি সংকেত বা তথ্য সংগ্রহ করার ক্ষমতাও নাটকীয়ভাবে বৃদ্ধি পায়। এই অগ্রগতিটা ধারনা করা হয় একটি নির্বাচনী চাপ বা সিলেক্টিভ প্রেশার তৈরী করে থাকতে পারে, যা কিনা ফটোরিসেপটর আর আইটপুট নিউরোনের সাদামাটা সংযোগ অপেক্ষা আরো বেশী উন্নত মানের রেটিনা ও সংকেত সম্পাদনার উদ্ভব তরান্বিত করতে সহায়তা করে। বিবর্তন প্রক্রিয়া এই প্রয়োজনটি মোকাবেলা করে রেটিনার অভ্যন্তরের কিছু কোষের পূর্ণতা পাবার প্রক্রিয়াটা খানিকটা পরিবর্তন করে, এর ফলে কিছু অপরিণত কোষ তাদের পুর্ণতা পাবার প্রক্রিয়ায় সিলিয়ারী রিসেপ্টর তৈরী করার বদলে রুপান্তরিত হয় রেটিনার বাইপোলার কোষে; যে কোষগুলো তাদের নিজেদের জায়গা করে নেয় ফটোরিসেপ্টর কোষ আর আউটপুট নিউরোনের মধ্যবর্তী স্তরে এই দুই ধরনের কোষের মধ্যে একটি সেতু বন্ধন তৈরী করে। এ কারনেই বিজ্ঞানীরা রেটিনার এই বাইপোলার কোষগুলোর সাথে এর আলোকসংবেদী কোষগুলো -রড এবং কোন – এর অনেক সাদৃশ্য খুজে পেয়েছেন। যদিও বাইপোলার কোষগুলোতে কোন ফটোপিগমেন্ট বা রোডাপসিন নেই, তারা আসলে আলোর বদলে ফটোরিসেপ্টরদের থেকে থেকে নি:সৃত রাসায়নিক পদার্থ (যাদের নিউরোট্রান্সমিটার বলা হয়) দ্বারা সক্রিয় হয়।
যদিও ক্যামেরা স্টাইল চোখ বেশ বিস্তৃত একটি দৃষ্টি সীমা দেয় (সাধারনতঃ ১৮০ ডিগ্রীর কাছাকাছি); কিন্তু ব্যবহারিক ক্ষেত্রে দেখা যায়, কোন একটি নির্দিষ্ট সময়ে আমাদের ব্রেন রেটিনা থেকে প্রাপ্ত মোট তথ্যের শুধুমাত্র খানিকটা বা অংশ বিশেষ একবারে প্রসেস করতে পারে। এই সীমাবদ্ধতার কারন, ব্রেনের সাথে আমাদের চোখের নিউরাল সংযোগ সীমিত কিছু স্নায়ু কোষের মাধ্যমে। একেবারের আদি ক্যামেরা স্টাইল চোখকে নিঃসন্দেহে আরো বেশী মাত্রার এ ধরনের সীমাবদ্ধতার মুখোমুখি হতে হয়েছে, কারন সম্ভবতঃ তখন স্নায়ু সংযোগকারী স্নায়ুকোষ বা নিউরনের সংখ্যা আরো কম ছিল। সুতরাং স্পষ্টত: প্রাকৃতিক নির্বাচনের যথেষ্ট চাপ ছিল চোখের নড়াচড়া নিয়ন্ত্রনের জন্য মাংশপেশীর বিবর্তনে। চোখের এই মাংশপেশীগুলোর নিশ্চয়ই উপস্থিত ছিল ৫০০ মিলিয়ন বছর আগে, এই ধারনার কারন হচ্ছে ল্যাম্প্রেদের, যাদের বংশধারা বা লিনিয়েজ এর বিস্তৃতি ছিল ঐ সময় পর্যন্ত্য, মধ্যে এদের উপস্থিতি আর বিন্যাস প্যাটার্ন,মানুষ সহ সকল চোয়ালযুক্ত মেরুদন্ডী প্রানীদের ক্ষেত্রে প্রায় হুবুহু এক রকম।
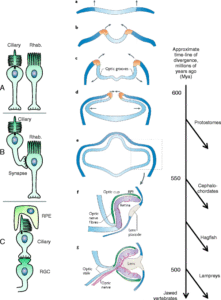
ছবি: মেরুদন্ডী প্রানীদের চোখের বিবর্তন প্রক্রিয়ার সিনারিও: বায়ে: সিলিয়ারী ও রাবডোমেরিক আলো সংবেদী কোষ: a) প্রোটোস্টোম ও ডিউটেরোস্টোম বিভাজনের আগে প্রায় ৫৮০ মিলিয়ন বছর আগে, আদি বাইল্যাটেরাল প্রানীদের দুই ধরনের: সিলিয়ারী ও রাবডোমেরিক আলোক সংবেদী কোষ ছিল। b) ডিউটেরোস্টোমস দের বিবর্তনের শুরুর দিকে ধারনা করা হয় রাবডোমেরিক ফটোরিসেপ্টরগুলো সিলিয়ারী রিসেপ্টরগুলো থেকে সংযোগ পেত : যা তখনো চোখের কেন্দ্রাভিমুখী হয়নি c) ধীরে ধীরে রাবডোমেরিক ফটোরিসেপ্টরগুলো তাদের ফটোরিসেপ্টর বৈশিষ্ট্যগুলো হারিয়ে ফেলে শুধু প্রজেকশন নিউরণ এ পরিনত (রেটিনাল গ্যাঙ্গলিয়ন সেল-RGC) হয়। সিলিয়ারী ফটোরিসেপ্টরগুলোর এই RGC দের সাথে সংযোগ থাকতেই হবে ভিটামিন এ র সঠিক আইসোমারটিকে পুণ সংশ্লেষন করার জন্য, যা ভিজুয়াল পিগমেন্ট অপসিনের জন্য জরুরী। এটা সম্ভব হয় আই ভেসিকলের ভিতর মুখী ভাজ খেয়ে যখন একটি আই ক্যাপ তৈরী করে ( ডান প্যানেলের f নং ছবি); মধ্যবর্তী ছবির প্যানেলে বর্তমানে জীবিত মেরুদন্ডী প্রানীদের ভ্রুণ বিকাশের সময় তাদের স্নায়ুতন্ত্রের ক্রমপরিবর্তন উপর ভিত্তি করে নিউরাল টিউব,আই ভেসিকল,আই ক্যাপ, লেন্স এর সম্ভাব্য বিবর্তনের ধারাবাহিকতা দেখানো হয়েছে। a থেকে d ) বিবর্তনের শুরুর দিকে বাইল্যাটেরাল প্রানীরা নিউরাল টিস্যুর ভিতরমুখী ভাজ বা ফোল্ডিং হয় যা কর্ডেটদের নিউরাল টিউব গঠন করে (৫৫০ মিলিয়ন বছর আগেই); ছবির কমলা রং নিউরাল টিউবের প্রান্তগুলোকে ইঙ্গিত করে; আলোক সংবেদী কোষগুলো এখানেই থাকে, যা দুই পাশে স্ফীত হয়ে আই ভেসাইকল তৈরী করে ( ড্যাশ দাগাঙ্কিত বক্স); ক্রমশ সম্প্রসারণশীল নিউরাল টিউব যেখানে ভ্রুণের বাইরের স্তর বা এক্টোডার্ম এর সংস্পর্শে আসলে তা অপটিক ভেসিকলে অন্তর্মুখী একটি ফোল্ড তৈরী প্রক্রিয়া শুরু করায় এবং ভেতরমুখী একটি আই ক্যাপের সৃষ্টি করে (f); যেখানে রেটিনা আর রেটিনাল পিগমেন্টাল এপিথেলিয়াম স্তর কাছাকাছি অবস্থান নেয়। ট্রেভর ল্যাম্ব ও তার সহযোগীদের প্রস্তাবনা হলো, এই আই ক্যাপ স্টেজটি (f) বিবর্তিত হয়েছে আমাদের লিনিয়েজ থেকে মিক্সিনয়েডদের লিনিয়েজ ( যাদের জীবিত প্রতিনিধি হলো হ্যাগফিশ) পৃথক হবার আগে। পরবর্তীতে যে লিনিয়েজ থেকে চোয়ালসহ এবং চোয়ালহীন মেরুদন্ড প্রানীদের জন্ম হয়েছে, তাদের এক্টোডার্ম ( বাইরের স্তর) পুরু বা মোটা হয়ে চোখের লেন্স গঠন করে (g)এই পর্বটি উদ্ভব হয়েছে যখন আমাদের আর ল্যাম্প্রেদের কমন আদি প্রানীদের উদ্ভবের সময়ে, সেটা প্রায় ৫০০ মিলিয়ন বছর আগে। ডানের প্যানেলে: একটি টাইমলাইন যা মাঝের প্যানেলের ধাপগুলোর বিবর্তনের একটি আপাত সময় নির্দেশ করে। সময়ের হিসাব ধরা হয়েছে মিলিয়ন বছর আগের হিসাব অনুযায়ী। সুত্র: ট্রেভর ল্যাম্ব ও সহযোগীরা : Evolution: Education and Outreach (1: 2008)
মেরুদন্ডী প্রানীদের চোখের মধ্যে বিবর্তন যত অসাধারন সুক্ষতম বৈশিষ্টই যোগ করুক না কেন, এর বেশ কিছু বড় মাপের ডিজাইন সমস্যাও আছে; যেমন আমাদের চোখের রেটিনাটা আসলে উল্টোভাবে সাজানো, অর্থাৎ ভেতরের দিকটা বাইরে আর বাইরের দিকটা ভেতরে। সুতরাং আলোকে রেটিনার আলোক সংবেদী পিগমেন্টের কাছে পৌছাতে রেটিনার পুরো প্রস্থটা পাড়ি দিতে হয়, যে পথে আছে স্নায়ুকোষের অসংখ্য প্রসেস দিয়ে তৈরী জালিকা, মধ্যবর্তী বাইপোলার কোষের স্তর, এছাড়া কোষ দেহ; যারা আলোকে বিভিন্ন দিকে ছড়িয়ে দিয়ে তাদের পথে প্রতবিন্ধকতা তৈরী করে এবং ফলে ইমেজ বা ছবির গুনগত মানও কমে যায়। অসংখ্য রক্তনালীকাও থাকে রেটিনার ভেতরের স্তরে, যারা অনাকাঙ্খিত ছায়া ফেলে রেটিনার ফটোরিসেপ্টর স্তরের উপর। এছাড়ার রেটিনার একটা ব্লাইন্ড স্পটও আছে (যেখানে কোন ফটোরিসেপ্টর কোষ থাকেনা), রেটিনার উপরের স্তর দিয়ে সাজানো বিস্তৃত নার্ভ ফাইবারগুলো এই ব্লাইন্ড স্পটে এসে একসাথে জড়ো হয়ে রেটিনার ভিতর দিয়ে একটি সুড়ঙ্গর মত পথ করে রেটিনা পেছন দিয়ে অপটিক নার্ভ হয়ে বের হয়ে যায়; এই ধরনের ডিজাইন সমস্যার তালিকা আরো দীর্ঘ।
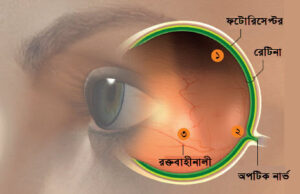
ছবি: বিবর্তনের চিহ্ন : মেরুদন্ডী প্রানীদের চোখ ভালো করলেই লক্ষ্য করা যায়, এটি আদৌ কোন ইন্টেলিজেন্ট ডিজাইনের নমুনা নয়, বরং এর মধ্যে অসংখ্য ক্রটি আছে, যা এর বিবর্তনের স্বাক্ষ্য বহন করে। এর কিছু ক্রটি আমাদের চোখের ইমেজ বা ছবি তৈরীর গুনগত মান কমিয়ে দেয়, যেমন তাদের মধ্যে আছে উল্টো করে সাজানো রেটিনা ( যার ভিতরের দিকটি বাইরে এবং বাইরের দিকটি ভিতরে), এ কারনে আলো কে একেবারে পেছনের স্তরে অবস্থিত ফটোরিসেপ্টরে পৌছাতে নানা ধরনের কোষ দেহ আর স্নায়ু ফাইবারের জালিকার মধ্যে দিয়ে যেতে হয়(১); রেটিনার উপর রক্তনালীকারা রেটিনার উপর অনাকাঙ্খিত ছায়া ফেলে (৩); সব স্নায়ু ফাইবারগুলো রেটিনা উপর দিয়ে এসে একজায়গায় জড়ো হয়ে রেটিনার মধ্য দিয়ে একটা ছিদ্র দিয়ে ঢুকে রেটিনার পিছনে অপটিক নার্ভ হিসাবে বের হয়ে যায়, এভাবে রেটিনার মধ্যে এটি একটি ব্লাইন্ড স্পট তৈরী করে (২); সুত্র: Illustration by Don Foley / Scientific American)
হয়তো প্রশ্ন আসতে পারে, ক্যামেরা স্টাইল কোন চোখের কি এগুলো এক ধরনের অপরিহার্য কোন সমস্যা ? উত্তর হচ্ছে, অবশ্যই না, এগুলো ক্যামেো স্টাইল চোখে আবশ্যিক কোন বৈশিষ্ট না। কারন অক্টোপাস আর স্কুইডেরও ক্যামেরা স্টাইল চোখ আছে যা বিবর্তিত হয়েছে মেরুদন্ডী প্রানী থেকে স্বতন্ত্র ভাবে, সেখানে কিন্ত এ ধরণের কোন ডিজাইন সমস্যা নেই। সৃষ্টিবাদী বা ইন্টেলিজেন্ট ডিজাইনের প্রস্তাবকরা যারা চোখকে একটি তিলোত্তমা অঙ্গ বলে দাবী করে আসছেন, এখানে তাদের নিজেদের যুক্তিতেই তারা পরাজিত হয়েছেন। আসলেই যদি কোন ইন্জ্ঞিনিয়ার আমাদের চোখের এই ডিজাইনের এই সব ভুল নিয়ে কোন একটা চোখ বানাতেন, তারতো চাকরীই থাকত না।
শধুমাত্র বিবর্তনের কাঠামোতে মেরুদন্ডী প্রানীদের চোখের বিবর্তন ব্যাখ্যা করলেই বোঝা সম্ভব আপাতঃ দৃষ্টিতে চোখের অদ্ভুত এইসব ক্রটিগুলো আসলে বিবর্তনের প্রাচীন পর্যায়গুলোরই ফলাফল, যার প্রত্যেকটি কোন না কোন ভাবে উপকৃত করেছে আমাদের প্রাচীন সব মেরুদন্ডী পুর্বসুরী প্রানীদের, এমনকি যখন তারা দেখতেই শুরু করেনি। আমাদের চোখের ডিজাইন অবশ্যই ইনটেলিজেন্ট ডিজাইন না, কিন্তু বিবর্তনের উজ্জ্বল আলোয় যখন একে পর্যবেক্ষন করা হয়, তখনই কেবল চোখের গঠন, এর পরিকল্পনা আর কাজটা স্পষ্ট করে বোঝা সম্ভবপর হয়।
ধন্যবাদ
প্রথম অংশ
চোখের বিবর্তন (The evolution of Eye) প্রথম অংশ



সাম্প্রতিক মন্তব্য